Сообщество дрозофил в ксерофитных растениях верхнего бассейна рек Парана и Парагвай
Машинный перевод
Оригинальная статья написана на языке EN (ссылка для ознакомления) .
Аннотация
Род Drosophila является самым многочисленным в семействе Drosophilidae. Некоторые виды являются эндемиками определенных регионов, а другие - космополитами. В Бразилии существует несколько экосистем для исследования состава и экологических аспектов Drosophila. Ксерофитные зоны являются примером. Их можно найти на юге и юго-востоке Бразилии в виде островов, что является результатом изменений в paleoclimatic циклах. Целью настоящей работы является предоставление информации о составе сообщества Drosophila в восьми ксерофитных зонах (где встречается кактус Cereus hildmaniannus ) на юге и юго-востоке Бразилии. Эта работа является важным шагом в изучении количественных экологических аспектов сообщества Drosophila в ксерофитных зонах, которые образуют континентальные острова в неотропическом регионе. Состав сообщества Drosophila , который был найден, совместим с предыдущими работами по нескольким аспектам. Экологические индексы показали возможную положительную ассоциацию между разнообразием и степенью сохранности изучаемых зон. Сообщества штата Сан-Паулу продемонстрировали наибольшее сходство среди протестированных участков, хотя статистически значимой корреляции между индексом Жаккара и географическим расстоянием не было найдено с использованием теста Мантела.
Введение
Хотя в 18 веке Скополи, Линней и Фабрициус первыми описали виды, которые сегодня относятся к семейству Drosophilidae, Дуда (1924a, 1924b, 1925) может считаться пионером в систематике этой группы. Более того, Стуртевант, хорошо известный своей классической работой (1921, 1939, 1942), является одним из самых важных исследователей в этом семействе. Первые данные о бразильском Drosophila сообществе были опубликованы Дудой (1925). В юго-восточном регионе Бразилии Добжанский и Паван (1943) были пионерами и описали несколько новых видов. После 1940 года было опубликовано много работ о бразильской фауне Drosophila (Паван и Кунья, 1947; Добжанский и Паван, 1950; Паван, 1950; 1959), включая первый список видов Drosophila Бразилии (Мурао и др., 1965). Позже другие авторы проанализировали фауну Drosophila в морфоклиматических доменах Бразилии в попытке лучше понять географическое распределение самых распространенных видов (Сене и др., 1980; Вилела и др., 1983; Валь и др., 1981).
Семейство Drosophilidae состоит из 65 родов и более 3,500 описанных видов, которые встречаются в различных экосистемах по всему миру (Bächli, 1998). Большинство родов встречаются в тропических регионах. Род Drosophila является самым многочисленным и составляет около 53% от общего числа видов. Некоторые из них являются эндемиками определенных регионов, а другие космополитичны, в основном распределены в связи с человеческой деятельностью. Гримальди (1990) рассмотрел классификацию семейства Drosophilidae на основе кладистического анализа с использованием максимальной парсимонии. Этот автор изучил 217 морфологических признаков взрослых особей и 120 видов, представляющих семью, и пришел к выводу, что они составляют монофилетическую группу. Род Drosophila вероятно возник в тропиках (Stalker, 1976), и различные его представители расширили свое распространение в умеренные зоны, где была отобрана способность использовать ресурсы в экстремальных экологических условиях. Этот процесс мог повторяться несколько раз в ходе эволюции рода.
Сегодня большинство исследований с использованием Drosophila проводятся в области молекулярной биологии и генетики, но только в начале двадцатого века Drosophila melanogaster была признана полезной для генетических анализов (Castle, 1906). Несмотря на большое количество исследований, использующих Drosophila в качестве модельного организма в этих областях, бразильские виды все еще требуют дальнейших исследований по многим аспектам экологии, систематики, генетики и эволюции.
Некоторые экологические исследования с Drosophila были проведены, особенно касающиеся активности мух в течение дня (Klaczko et al., 1983; Medeiros, 2000), подходящего времени суток для отбора проб (Pavan et al., 1950; Belo & Oliveira-Filho, 1978) и пространственного распределения видов (горизонтальное: Dobzhanzky & Pavan, 1950; Burla et al., 1950; Pavan, 1959; Medeiros, 2000; и вертикальное: Sene et al., 1981; Kratz et al., 1982 и Tidon-Sklorz & Sene, 1992). Однако сравнение между районами получило мало внимания (Medeiros & Klaczko, 2004, например) и по-прежнему остается открытой областью исследования для бразильских видов Drosophila.
Учитывая это, в Бразилии есть несколько экосистем для изучения в отношении состава и экологических аспектов Drosophila. Ксерофитные районы, определяемые наличием растительности сухого типа с различными видами кактусов, являются таким примером. В настоящее время, а также в центральных районах произрастания кактусов, таких как бразильская Катынга и аргентинский Чако, эти растения можно найти на юге, юго-востоке и центральных регионах Бразилии на островах, что является результатом циклических палеоклиматических изменений. Эти циклы чередовались между холодными/сухими и теплыми/влажными. В первом случае ксерофитные районы расширялись из центральных областей по континенту, а во втором они сокращались, образуя убежища (острова) между ними (Бигарелла и др., 1975; Аб’Сабер, 1977; Ванзоли, 1981). Таким образом, убежища важны для биоразнообразия, экологических и эволюционных исследований нескольких групп, включая виды Drosophila, которые размножаются специально в гниющей ткани кактусов, как и мухи группы D. repleta.
Цель настоящей работы заключается в предоставлении информации о составе сообщества Drosophila в восьми ксерофитных районах (где встречается кактус Cereus hildmaniannus) в южных и юго-восточных регионах Бразилии. Это предоставит ценную информацию для дальнейших исследований в области экологии, генетики и эволюции географически изолированных популяций Drosophila.
Материалы и методы
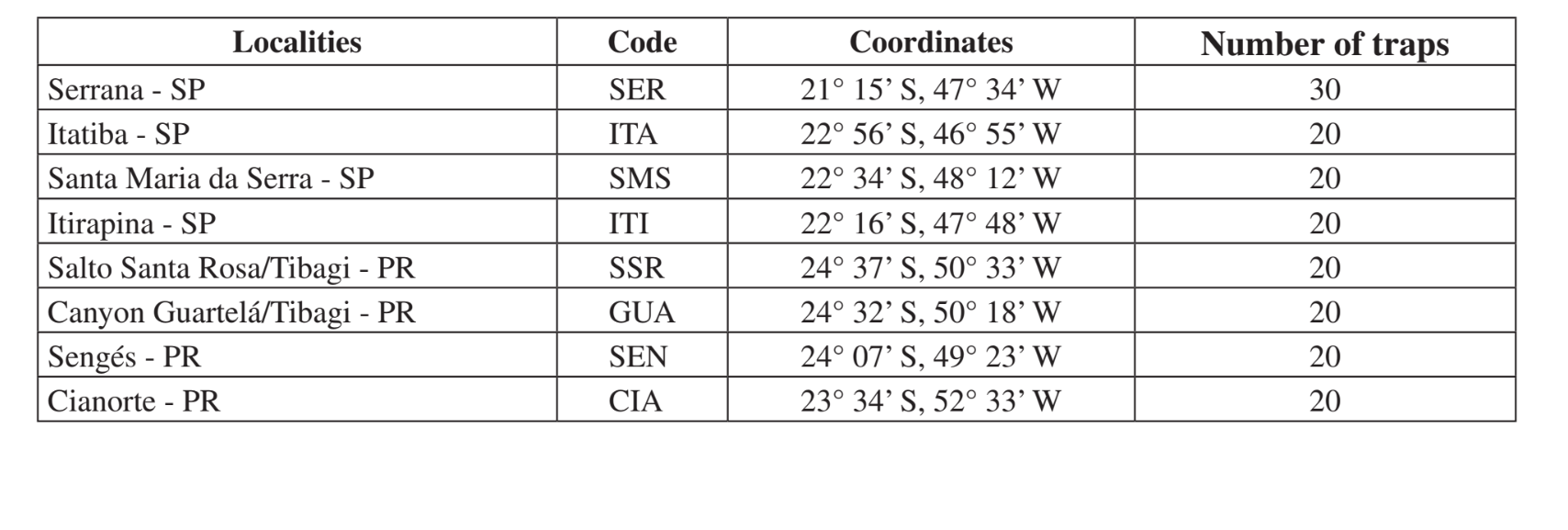
Места и даты сборов суммированы в Таблице 1. Образцы были собраны в лесах, состоящих из ксерофитной растительности в мезофитных лесах в долинах верхней части бассейна рек Парана-Парагвай. Эти районы находятся на разных стадиях нарушения, начиная от использования для разведения скота рядом с дорогой (Итатиба - SP и Сенжес - PR) до зоны охраны Государственного парка (Каньон Гуартелá - PR). Все сборы были проведены в конце влажного периода, который соответствует февралю-майю в два разных года (1999 и 2000).
Сборы
В течение трех дней взрослые мухи привлекались к ферментированным бананам и апельсинам в открытых ловушках, расположенных не ближе 15 м друг от друга, висящих на ветвях на высоте ± 1,5 м от земли. После этого периода мухи были пойманы энтомологическими сетями, помещены в стеклянные банки с культурной средой и доставлены в лабораторию живыми, где они были идентифицированы.
Определить представителей групп Drosophila repleta и D. willistoni до определенной степени сложно. Самки первой группы считаются неопределенными, если диагностическим признаком является адеагус самца. В группе Drosophila willistoni идентификация с использованием генетических признаков более надежна, но не проводилась. Поэтому криптические виды были сгруппированы как “repleta группа” и “D. willistoni группа”, соответственно. Неопределенные виды, похожие на “repleta группу”, не учитывались в анализах.
Анализ данных
Для качественной проверки появления видов использовался Метод Постоянства Появления (Dajoz, 1983). Значение постоянства (c) получалось путем деления числа коллекций, в которых встречался один вид, на общее число коллекций, а затем умножением этого результата на 100. Виды с индексом c ≥ 50 считались постоянными. Вспомогательные виды были теми, у которых 25 ≤ c < 50. Случайные виды имели c < 25. Виды, встречающиеся только в одной области, считались эксклюзивными.
Drosophila сообщества анализировались с использованием нескольких экологических индексов: Бергера-Паркера (1/d), Шеннона-Уинера (H’), Симпсона (D), Маргалефа (DMg), Менхиника (DMn) и Жаккара (J). Для определения различий в медианной abundancy между сообществами использовалась непараметрическая ANOVA, Краскала-Уоллиса. Затем проводилось множественное сравнение между образцами (аналогично тесту Тьюки), чтобы определить, произошли ли значительные различия между образцами (Zar, 1999).
Для проверки взаимосвязей между сообществами были проведены два кластерных анализа:
1) с использованием присутствия и отсутствия видов; и 2) с использованием количества собранных экземпляров для каждого вида в каждой области. Фенограммы были построены в программном обеспечении Minitab® для Windows Release 10.1, выбрав методы евклидова расстояния и одиночной связи в обоих подходах.
Чтобы проанализировать, существует ли ассоциация между сходствами сообществ и географическими расстояниями, был проведен тест Мантела с использованием программного обеспечения TFPGA (Miller, 1997), сравнивающий две матрицы: индексы Жаккара и географические расстояния между парами сообществ.
Результаты и обсуждение
В этом исследовании мы смогли идентифицировать в общей сложности 10,464 мухи, принадлежащие к 4 подроду, 10 группам и как минимум 30 различным видам Drosophila (Таблица 2). Постоянные виды (c ≥ 50) составили примерно 63% от общего числа собранных видов (19 из 30). Семь видов были признаны вспомогательными (~ 23%), и было найдено 4 случайных вида (~ 13%). Все подроды показали постоянные виды, за исключением подрода Dorsilopha , который показал только один случайный вид (Drosophila busckii).
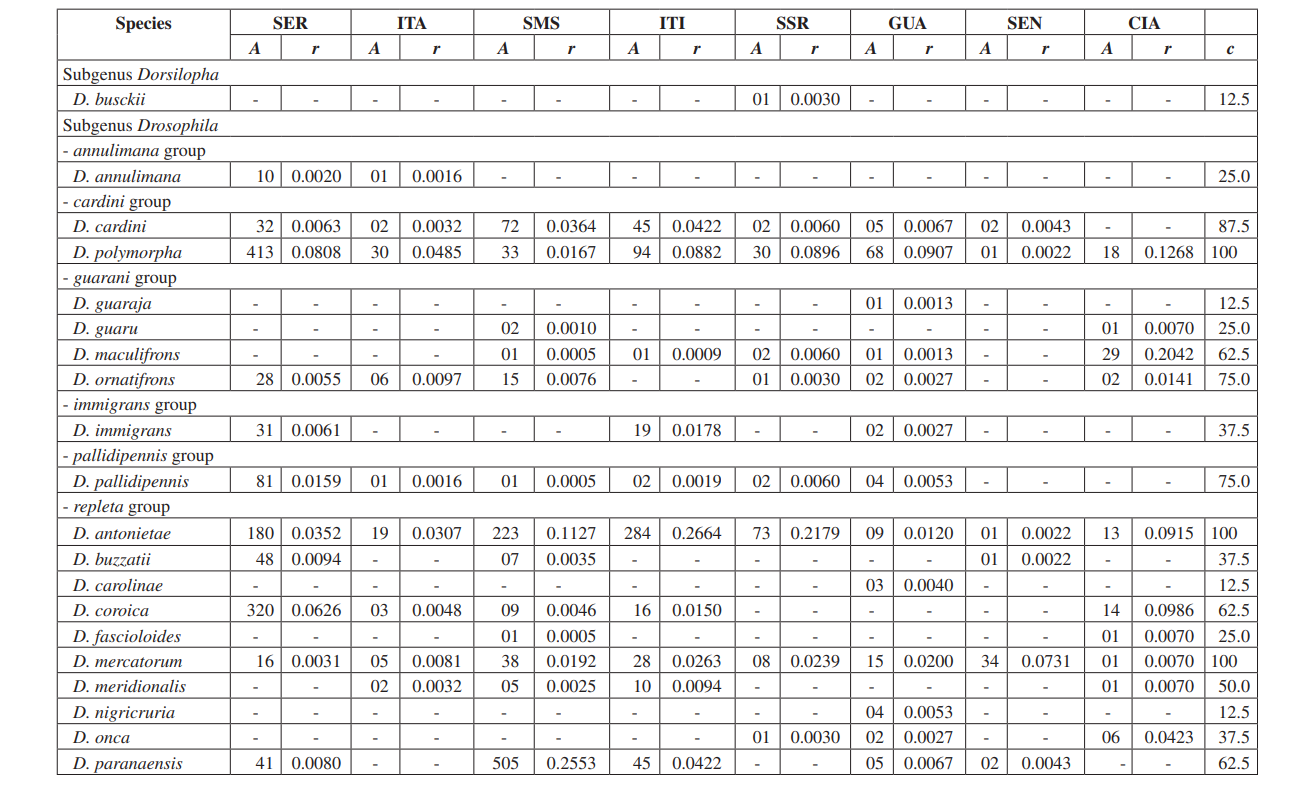
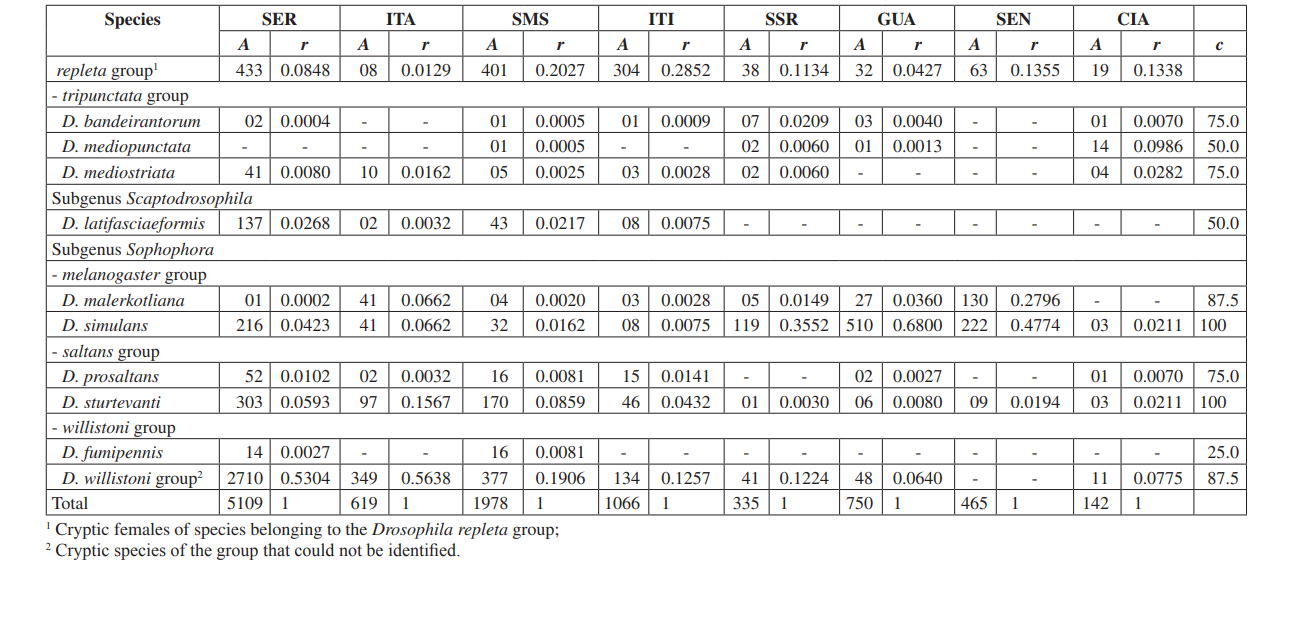
Пять видов (Drosophila polymorpha, D. antonietae, D. mercatorum, D. simulans и D. sturtevanti) были признаны многочисленными и были собраны во всех 8 районах (Серрана - SP, Итатиба - SP, Санта-Мария да Серра - SP, Итирапина - SP, Сальто Санта-Роза - PR, Каньон Гуартелá - PR, Сенгеш - PR и Сианорте - PR). Этот результат соответствует предыдущим обследованиям Drosophila.
Drosophila polymorpha была собрана от Гватемалы до Бразилии и в различных типах сред: лесах, серраду, рестинге, а также в ассоциации с человеческим присутствием. Это относительно многочисленный вид в различных морфоклиматических доменах, за исключением каатинга (Сене и др., 1980). Drosophila antonietae ожидалась в сборе во всех районах, так как это кактусофильный вид, который встречается на юге и юго-востоке Бразилии, а также на севере восточной границы аргентинского Чако. Этот вид всегда встречается в ассоциации с колонновидным кактусом Cereus hildmaniannus в ксерофитной растительности галерейных и мезофитных лесов в долинах бассейна реки Парана-Уругвай (Тидон-Склорз & Сене, 2001). Этот паттерн распределения, который изолирует Drosophila antonietae от других видов кластера D. buzzatii, совпадает с определенными паттернами других двукрылых и также с регионами эндемизма, что предполагает викариантные события, такие как зоогеографические барьеры, установленные в более ранние геологические эпохи, или климатические события, которые препятствовали генетическому потоку, позволяя накапливаться различиям между популяциями (Аморин & Пирес, 1996). Этот паттерн распределения также наблюдается в отношении распределения растений, ограниченного ксерофитным каатингом и регионами чако (Дювернель & Инес, 2000).
Drosophila mercatorum довольно распространен в естественных условиях в Южной Америке, особенно в открытых местах (Sene и др., 1981; Vilela и др., 1983).
Drosophila simulans является интродуцированным видом, который был собран в большом количестве в нескольких местах (Dobzhansky & Pavan, 1950; Pavan, 1959; Sene и др., 1980; Tidon-Sklorz & Sene, 1992), также в открытых местах (Perondini и др., 1979).
Drosophila sturtevanti является широко распространенным видом в Центральной и Южной Америке, встречающимся в различных морфоклиматических доменах (Tidon-Sklorz & Sene, 1999).
Эксклюзивные виды были обнаружены в Salto Santa Rosa - PR (Drosophila busckii) и Canyon Guartelá - PR (Drosophila guaraja, D. carolinae и D. nigricruria). Drosophila busckii не ожидали собрать в Salto Santa Rosa - PR, так как это интродуцированный вид в Бразилии и часто ассоциируется с измененными человеком средами (Tidon-Sklorz & Sene, 1999). Тем не менее, его собирали в естественных условиях, в основном в сухих открытых растениях (Bizzo & Sene, 1982; Tidon-Sklorz & Sene, 1992). Среди эксклюзивных видов, собранных в Canyon Guartelá - PR, Drosophila guaraja был зарегистрирован в Бразилии только в Boracéia - SP (Val & Kaneshiro, 1988). Таким образом, это первая запись о нем за пределами этой местности в Бразилии. Drosophila carolinae был собран только в тропических лесах в Бразилии (Vilela, 1983) и по этой причине не ожидали найти его в районах ксерофитной растительности. Тем не менее, Canyon Guartelá - PR является одной из наиболее сохранившихся обследованных территорий, и кактусы распределены в лесной зоне внутри Государственного парка, что может объяснить его присутствие. Drosophila nigricruria является единственным эксклюзивным видом с широкой географической распространенностью. Тем не менее, его собрали только в этой области в данном исследовании. Medeiros & Klaczko (2004) также собрали этот вид на ферме Barreiro Rico (между 48° 05’ 11’’ W – 22° 41’ 15’’ S и 48° 04’ 52’’ W – 22° 41’ 26’’ S), на внутреннем плато, где находится полудекоративный лес и один из трех оставшихся лесов в штате Сан-Паулу, где были собраны образцы этими авторами.
Подрод Drosophila показал наибольшее разнообразие (7 групп и 22 вида), из которых 7 видов принадлежат группе Drosophila repleta . Второе по величине разнообразие было обнаружено в подроде Sophophora (3 группы и 6 видов). Это распределение разнообразия среди подродов было отмечено в других исследованиях (Sene и др., 1981; Tidon-Sklorz & Sene, 1995; Tidon-Sklorz & Sene, 1999; Medeiros & Klaczko, 2004).
Введенный вид дрозофил, Zaprionus indianus (Vilela, 1999), был найден во всех коллекциях (1,459 образцов в Серране; 317 в Итатибе; 409 в Санта-Марии да Серра; 302 в Итирапине; 42 в Салто Санта Роза; 117 в Гуартелá; 598 в Сенгеше; и 8 в Сианорте). События, связанные с биологическими инвазиями, делятся на три категории: прибытие, установление и интеграция (Vermeij, 1996). Согласно Moraes и др. (2000), Zaprionus indianus прошел через все три стадии и, вероятно, конкурирует неблагоприятно с другими схожими видами на юго-востоке Бразилии, так как было отмечено снижение относительной частоты. Введение экзотического вида Drosophila в Бразилии не является редким событием. В 1976 году Val & Sene (1980) собрали большое количество Drosophila malerkotliana на северо-востоке Бразилии, вид, который не был зафиксирован в обширных предыдущих коллекциях. В настоящее время в наших обследованиях Drosophila malerkotliana собирается почти по всему Бразилии, хотя всегда с низкой относительной частотой.
В Санта-Мария да Серра - SP было собрано двадцать три различных вида, 20 видов в Серране - SP и в Каньоне Гуартелá - PR, 18 в Итирапине - SP, 17 в Сианорте - PR, 16 в Итатибе - SP и Сальто Санта Роза - PR и 9 в Сенгеше - PR (Таблица 3). Медеирос и Клачко (2004) описали последний инвентаризационный отчет о фауне Drosophila в Бразилии, проведенный в трех оставшихся лесах в штате Сан-Паулу. Эти авторы получили 57, 76 и 90 различных видов в трех изученных областях. Эти числа видов больше, чем наблюдаемые здесь, возможно, в результате различий в методах сбора или просто потому, что обследованные области отличаются по своему растительному составу (лес и ксерофитная растительность). Наибольшее общее количество было обнаружено в Серране - SP (4676 экземпляров), а наименьшее в Сианорте - PR (123 экземпляра) (Таблица 3).
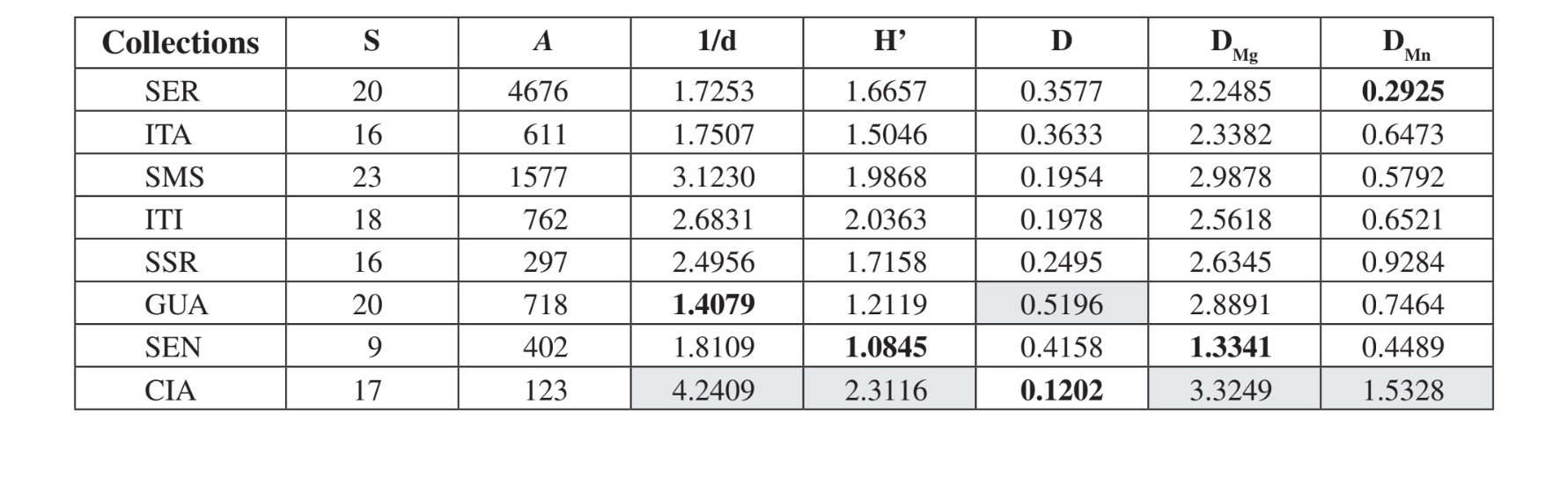
Согласно экологическим индексам (Таблица 3), Сианорте - ПР показал более высокие значения почти по всем индексам (1/d = 4.2409; H’ = 2.3116; DMg = 3.3249; DMn = 1.5328), за исключением Симпсона (D), который показал, что Каньон Гуартелá - ПР является самой
разнообразной областью (D = 0.5196). Каньон Гуартелá - ПР также показал один из самых высоких индексов в двух других случаях (DMg = 2.8891; DMn = 0.7464). Сенгеш - ПР, Итатиба - СП и Серрана - СП показали самые низкие значения по большинству индексов. Санта-Мария да Серра - СП, Итирапина - СП и Сальто Санта Роза - ПР показали промежуточные значения по большинству индексов. Эти наблюдения соответствуют поверхностной оценке деградации в этих районах. Сианорте - ПР расположен рядом с дорогой, но на высоком склоне реки Лижейру, с трудным доступом для человеческой деградации. Каньон Гуартелá - ПР является лесной зоной внутри Государственного парка Каньон Гуартелá в штате Парана, как уже упоминалось ранее. Поэтому это высоко охраняемая территория. Сенгеш - ПР является самой деградированной областью, рядом с дорогой, где ведется скотоводство и под высоким влиянием человеческой деятельности. Серрана - СП - это охраняемая территория на вершине холма, к которой трудно добраться. Более низкие индексы разнообразия, найденные для этой области, должны быть связаны с большим количеством ловушек, использованных в этой коллекции (Таблица 1). Ожидалось, что другие области будут иметь промежуточные значения разнообразия, так как они являются небольшими охраняемыми территориями внутри ферм.
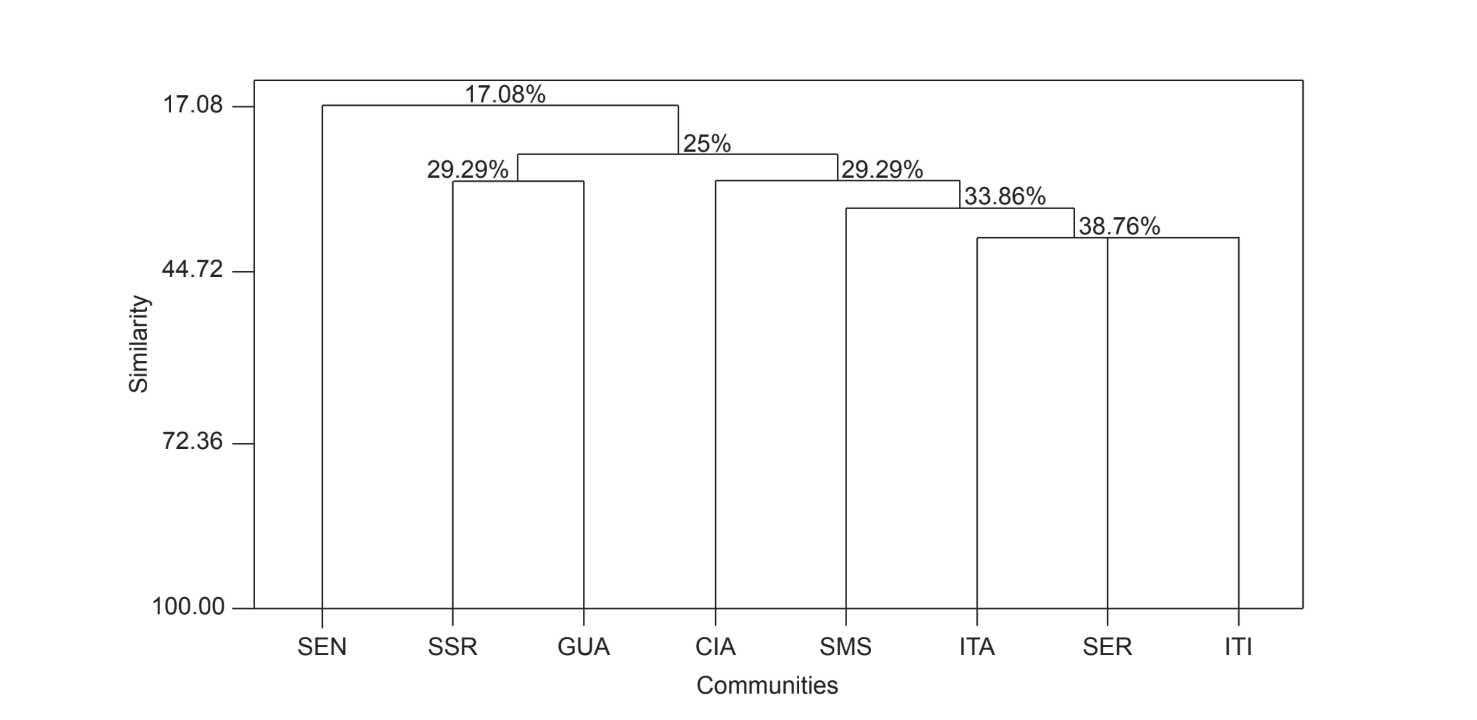
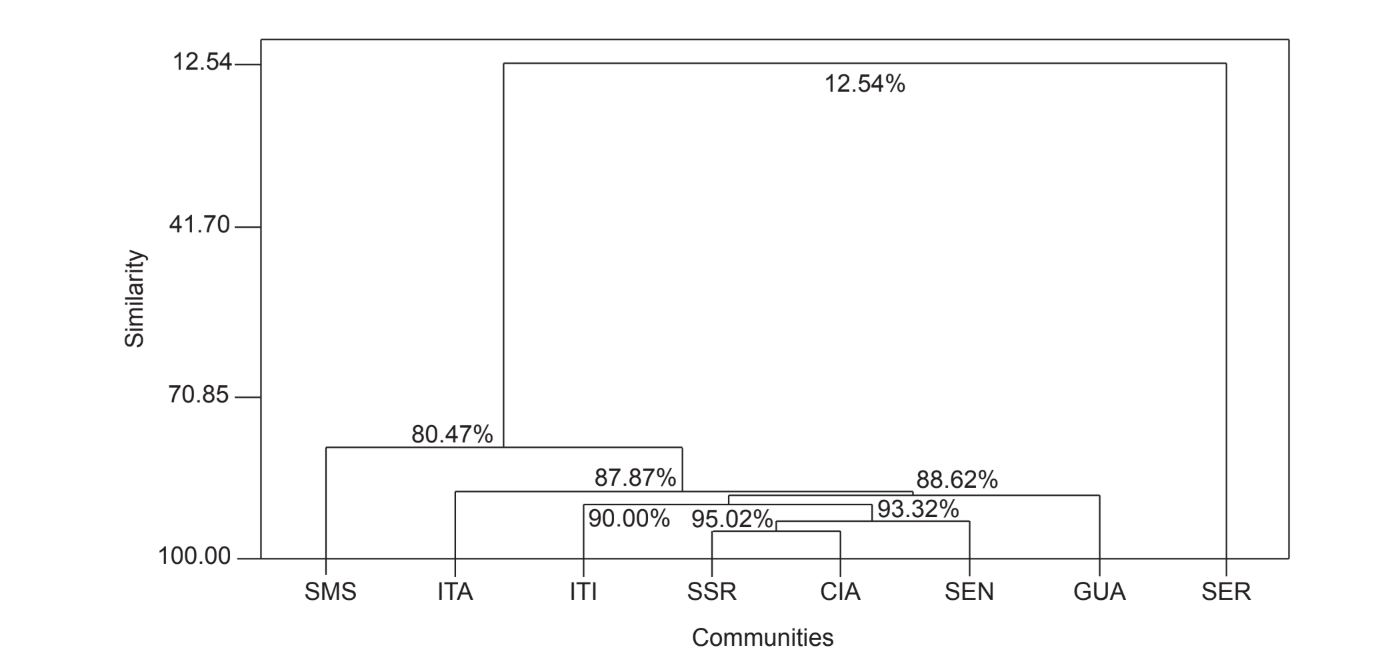
Парное сравнение сообществ с использованием индекса Жаккара (Таблица 4) показало, что сообщества штата Сан-Паулу (Серрана - SP, Итатиба - SP, Санта-Мария да Серра - SP и Итирапина - SP) имели наивысшую схожесть среди всех сравнений (J > 0.62). Тот же паттерн не наблюдался среди сообществ штата Парана, а также между обоими штатами, за исключением индекса, полученного между Санта-Мария да Серра - SP и Сианорт - PR (J = 0.667), и между Сальто Санта Роза - PR и Каньоном Гуартелá - PR (J = 0.636). Кластерный анализ с использованием данных о присутствии и отсутствии видов (Рис. 1), несмотря на низкий уровень схожести (33.86%), также объединил все сообщества штата Сан-Паулу. Наиболее отличающимся сообществом был Сенгеш - PR. Рис. 2 изображает кластерное объединение всех сообществ с использованием данных об абсолютной численности (A). Были обнаружены более высокие уровни схожести, но четкой ассоциации сообществ не было. На этом рисунке Серрана - SP была наиболее отличающимся сообществом.
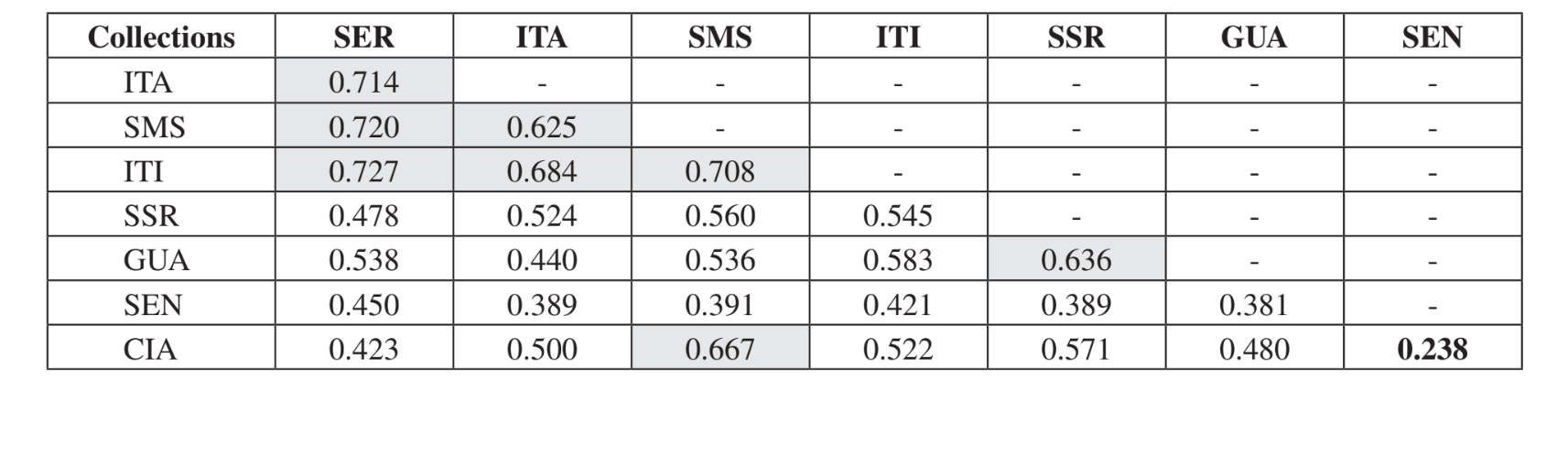
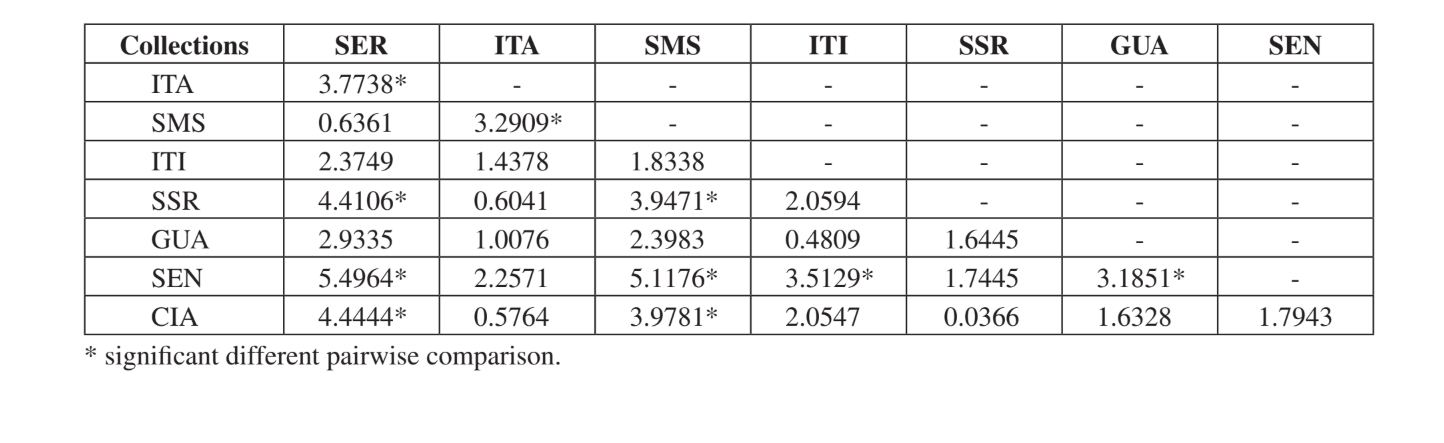
Тест Крускала-Уоллиса, скорректированный на совпадения, показал, что сообщества значительно различаются при общем сравнении (H = 24.24; d.f. = 7; p = 0.001). Множественные парные сравнения сообществ представлены в Таблице 5. Сообщества Серрана - SP, Санта-Мария да Серра - SP и Сенгеш - PR были наиболее отличительными при индивидуальном сравнении с другими (по 4 значимых различия для каждого). Итирапина - SP и Каньон Гуартелá - PR были наиболее схожими сообществами при сравнении с каждым из исследованных сообществ (только одно значимое различие для каждого). Все эти сравнения (индекс Жаккара, кластерный анализ и множественные парные сравнения) показали, что более высокая схожесть наблюдалась среди сообществ штата Сан-Паулу, хотя положительная значимая корреляция не была найдена в тесте Мантела, сравнивающем индекс Жаккара и географические расстояния (r = – 0.3235; p = 0.0839).
Опрос о составе сообщества является важным этапом для нескольких исследований в широком диапазоне областей, таких как генетика, экология и эволюция. Эта работа добавляет новые знания о составе и разнообразии сообщества Drosophila в ксерофитных районах, которые образуют континентальные острова в неотропическом регионе. Этот подход важен для дальнейших исследований в любой области, рассматривающей Drosophila как модель.
Авторы: Матейш, Р. П., Бушини, М. Л. Т. и Сене, Ф. М
Ссылки:
- Аб’сабер, А. Н., 1977, Пространства, занятые расширением сухих климатов Южной Америки, во время ледниковых периодов четвертичного периода. Палеоклиматы, 3: 1-19.
- Аморин, Д. С. и Пирес, М. Р. С., 1996, Неотропическая биогеография и метод максимальной оценки биоразнообразия. В: К. Е. М. Бикудо и Н. А. Менезес (ред.), Биоразнообразие в Бразилии, первый подход, CNPq, Сан-Паулу.
- Бахли, Г., 1998, Семейство Drosophilidae. В: Л. Папп и Б. Дарвас (ред.), Вклад в руководство по палеарктическим двукрылым. III. Высшие брахицеры, Научный вестник, Будапешт.
- Белу, М. и Оливейра-Фильо, Ж. Ж., 1978, Домашние виды Drosophila. V. Влияние экологических факторов на количество пойманных особей. Rev. Bras. Biol., 36: 903-909.
- Бигарелла, Ж. Ж., Андраде-Лима, Д. и Риес, П. Ж., 1975, Соображения относительно палеоклиматических изменений в распределении некоторых растительных и животных видов в Бразилии. An. Acad. Brasil. Ciênc., 41: 411-464.
- Биццо, Н. М. В. и Сене, Ф. М., 1982, Исследования естественных популяций Drosophila из Перуибе (SP), Бразилия (Двукрылые, Drosophilidae). Rev. Bras. Biol., 42: 539-544.
- Бурла, Х., да Кунья, А. Б., Кавалканти, А. Г. Л., Паван, К. и Добжанский, Т., 1950, Плотность населения и скорость расселения в бразильской Drosophila willistoni. Экология, 31: 393-404.
- Кастл, У. Е., 1906, Инбридинг, скрещивание и стерильность в Drosophila. Наука, 23: 153.
- Дажоз, Р., 1983, Общая экология. 471 стр. Издательство Возес, Петрополис.
- Добжанский, Т. и Паван, К., 1943, Исследования бразильских видов Drosophila. Bolm. Fac. Filos. Ciênc. S. Paulo, 36 (Biol. Geral, 4): 1-72.
- Добжанский, Т. и Паван, К., 1950, Местные и сезонные вариации в относительных частотах видов Drosophila в Бразилии. J. Anim. Ecol., 19: 1-14.
- Дуда, О., 1924a, Вклад в систематику Drosophilidae с особым учетом палеарктических и ориентальных видов (Двукрылые). Arch. Naturgesch., 90: 172-234.
- Дуда, О., 1924b, Ревизия европейских видов рода Drosophila Fallén (Двукрылые). Ent. Medd., 14: 246-313.
- Дуда, О., 1925, Костариканские Drosophilidae Венгерского национального музея в Будапеште. Annls hist.- nat. Mus. natn. hung., 22: 149-229.
- Дювернель, Д. Д. и Иэйнс, У. Ф., 2000, Контрастирующая молекулярная популяционная генетика четырех гексокиназ в Drosophila melanogaster, D. simulans и D. yakuba. Генетика, 156: 1191-1201.
- Гримальди, Д. А., 1990, Филогенетическая пересмотренная классификация родов в Drosophilidae (Двукрылые). Bull. Am. Mus. nat. Hist., 197: 1-139.
- Клачко, Л. Б., Пауэлл, Дж. Р. и Тейлор, С. Е., 1983, Drosophila приманки и дрожжи: привлекаемые виды. Оэколоджия, 59: 411-413.
- Кратц, Ф. Л., Пинто, Л. Г., Брандано, Д. и Фария, Л. Г., 1982, Высота полета и пространственное распределение в Drosophila. Ciênc. Cult., 34: 203-209.
- Медейрос, Х. Ф., 2000, Ассамблеи видов Drosophila (Двукрылые; Drosophilidae) и влияние водных потоков на их распределение в двух лесах Сан-Паулу, 98 стр. Диссертация (магистратура) – Институт биологии IB/UNICAMP, Кампинас.
- Медейрос, Х. Ф. и Клачко, Л. Б., 2004, Сколько видов Drosophila (Двукрылые, Drosophilidae) еще предстоит описать в лесах Сан-Паулу, Бразилия? Списки видов трех лесных остатков. Biota Neotropica, 4: 1-12.
- Миллер, М. П., 1997, Инструменты для анализа популяционной генетики - TFPGA - 1.3: Программа для Windows для анализа аллозимных и молекулярных данных популяционной генетики. Компьютерное программное обеспечение, распространяемое автором.
- Мораес, Е. М., Матейш, Р. П. и Сене, Ф. М., 2000, Происхождение и колебания в численности популяции недавно введенного вида Drosophilidae в Бразилии. В: АННОТАЦИИ XXI МЕЖДУНАРОДНОГО КОНГРЕССА ЭНТОМОЛОГИИ, КНИГА I. Материалы..., 271 стр.
- Мурао, К. А., Галло, А. Ж. и Бикудо, Х. Е. М. С., 1965, О систематике Drosophila в Бразилии, с описанием D. mendeli sp.n. и "Список бразильских видов рода Drosophila". Ciênc. Cult., 17: 577-585.
- Паван, К., 1950, Бразильские виды Drosophila. II. Bolm. Fac. Filos. Ciênc. S. Paulo, 111 (Biol. Geral, 8): 1-37.
- Паван, К., 1959, Связи между естественными популяциями Drosophila и окружающей средой. Bolm. Fac. Filos. Ciênc. S. Paulo, 221 (Biol. Geral, 11): 1-81.
- Паван, К. и да Кунья, А. Б., 1947, Бразильские виды Drosophila. Bolm. Fac. Filos. Ciênc. S. Paulo, 86 (Biol. Geral, 7): 3-47.
- Паван, К., Добжанский, Т. и Бурла, Х., 1950, Дневное поведение некоторых неотропических видов Drosophila. Экология, 31: 36-43.
- Перондини, А. Л. П., Сене, Ф. М. и Мори, Л., 1979, Шаблон и полиморфизм некоторых Drosophila simulans эстераз в Бразилии. Egypt. J. Genet. Cytol., 8: 263-268.
- Сене, Ф. М., Валь, Ф. С., Вилела, К. Р. и Перейра, М. А. Q. Р., 1980, Предварительные данные о географическом распределении видов Drosophila в пределах морфоклиматических доменов Бразилии. Pap. Avul. Dep. Zool. Sec. Agric., 33: 315-326.
- Сене, Ф. М., Перейра, М. А. Q. Р., Вилела, К. Р. и Биццо, Н. М. В., 1981, Влияние различных способов установки приманок для сбора мух Drosophila в трех естественных средах. D. I. S., 56: 118-121..
- Стаклер, Х. Д., 1976, Исследования хромосом в дикой популяции D. melanogaster. Генетика, 82: 323-347.
- Стуртевант, А. Х., 1921, Североамериканские виды Drosophila. Carnegie Inst. Wash. Publ., 301: 1-150.
- Стуртевант, А. Х., 1939, О подразделении рода Drosophila. Proc. Natl. Acad. Sci. USA, 3: 137-141.
- Стуртевант, А. Х., 1942, Классификация рода Drosophila, с описанием девяти новых видов. Univ. Texas Publ., 4213: 6-51.
- Тидон-Склорц, Р. и Сене, Ф. М., 1992, Вертикальное и временное распределение видов Drosophila (Двукрылые, Drosophilidae) в лесистой местности в штате Сан-Паулу, Бразилия. Rev. Bras. Biol., 52: 311-317.
- Тидон-Склорц, Р. и Сене, Ф. М., 1995, Фауна Drosophila (Двукрылые, Drosophilidae) в северной части "Кадеиа до Эспиньясу", штаты Минас-Жерайс и Баия, Бразилия: биогеографические и экологические аспекты. Iheringia, Sér. Zool., 78: 85-94.
- Тидон-Склорц, Р. и Сене, Ф. М., 1999, Drosophila. В: К. Р. Ф. Брандано и Е. М. Канселло (ред.), Наземные беспозвоночные. Т. V. Биоразнообразие штата Сан-Паулу. Синтез знаний к концу XX века, FAPESP, Сан-Паулу.
- Тидон-Склорц, Р. и Сене, Ф. М., 2001, Два новых вида из группы сиблингов Drosophila serido (Двукрылые, Drosophilidae). Iheringia, Sér. Zool., 90: 141-146.
- Валь, Ф. С. и Канеширо, К. Й., 1988, Drosophilidae (Двукрылые) из Биологической станции Боросейя, на прибрежном хребте штата Сан-Паулу, Бразилия: Географическое распределение. В: П. Е. Ванзолини и У. Р. Хейер (ред.), Материалы семинара по неотропическим паттернам распределения, Бразильская академия наук, Рио-де-Жанейро.
- Валь, Ф. С. и Сене, Ф. М., 1980, Новый введенный вид Drosophila в Бразилии. Pap. Avul. Dep. Zool. Sec. Agric., 33: 293-298.
- Валь, Ф. С., Вилела, К. Р. и Маркес, М. Д., 1981, Drosophilidae неотропического региона. В: М. Эшбернер, Х. Л. Карсон и Дж. Н. Томпсон (ред.), Генетика и биология Drosophila, Академическое издательство, Лондон.
- Ванзолини, П. Е., 1981, Квази-исторический подход к естественной истории дифференциации рептилий в тропических географических изолятах. Pap. Avul. Dep. Zool. Sec. Agric., 34: 189-204
- Вермеий, Г. Дж., 1996, Повестка дня для биологии инвазий. Биол. Консервация, 78: 3-9.
- Вилела, К. Р., 1983, Ревизия группы видов Drosophila repleta (Двукрылые, Drosophilidae). Rev. Bras. Entomol., 27: 114.
- Вилела, К. Р., 1999, Является ли Zaprionus indianus Гупта, 1970, (Двукрылые, Drosophilidae) в настоящее время колонизирующим неотропический регион? D. I. S., 82: 37-39.
- Вилела, К. Р., Перейра, М. А. Q. Р. и Сене, Ф. М., 1983, Предварительные данные о географическом распределении видов Drosophila в пределах морфоклиматических доменов Бразилии. II. Группа repleta. Ciênc. Cult., 35: 66-70.
- Зар, Дж. Х., 1999, Множественные сравнения. В: Биостатистический анализ. 4-е изд., Prentice Hall Inc., Нью-Джерси.