Характеризация брачных звуков видов подсубгруппы Fasciola (Двукрылые, Drosophilidae, группа Drosophila repleta): межвидовые и межпопуляционные анализы
Машинный перевод
Оригинальная статья написана на языке EN (ссылка для ознакомления) .
Аннотация
Целью данной работы было охарактеризовать паттерн мужской брачной песни различных видов подгруппы fasciola и определить уровень вариации как внутри видов, так и между ними. Анализировались параметры: интервал внутри импульса (PI), интервал между импульсами (IPI) и частота внутри импульса (IF). Были проанализированы шесть различных видов: D. coroica (три популяции), D. ellisoni, D. fascioloides, D. moju, D. onca и D. rosinae (по одной популяции каждого). Были выявлены значительные различия между шестью видами по этим трем параметрам брачной песни. IPI оказался самым изменчивым параметром среди этих видов, что предполагает, что этот параметр важен для дискриминации самок. Четыре различные гипотезы могут объяснить эту вариацию: 1. различные селекционные давления при отсутствии генетического потока; 2. интраспецифическая сексуальная селекция; 3. симпатрические эффекты на эволюцию песни; и 4. генетический дрейф. PI был единственным параметром, который значительно отличался среди трех популяций D. coroica. Низкая изменчивость среди популяций внутри одного вида уже была замечена для других подгрупп и может быть объяснена следующими гипотезами: сильный отбор, действующий на параметры песни, генетический поток или недавняя колонизация из общего источника. Дополнительные исследования брачной песни других видов подгруппы fasciola, а также для других подгрупп группы repleta, и исследования с использованием молекулярных маркеров, которые сосредоточены на генетической основе различий между этими видами в брачной песне, позволят нам оценить связь между брачной песней и сексуальной изоляцией у этих видов, а также помогут понять эволюцию этих поведенческих различий.
Введение
Звуки ухаживания
Ухаживающее поведение мух рода Drosophila важно для распознавания видов. После того как Шорей (1962) показал, что самцы D. melanogaster издают звуки во время сексуального ухаживания, было проведено множество исследований с различными группами и подгруппами рода Drosophila, описывающими специфическую природу типов звуков. Эти исследования предоставили доказательства того, что звук выполняет функцию как сексуального стимула, так и в сексуальной изоляции. Эти звуки значительно различаются среди различных видов группы affinis (Chang & Miller, 1978), melanogaster (Cowling & Burnet, 1981), virilis (Hoikkala et al., 1982; Hoikkala & Isoherranen, 1997), mercatorum (Ikeda & Maruo, 1982), repleta (Ewing & Miyan, 1986), auraria (Tomaru & Oguma, 1994), planitibia (Hoikkala et al., 1994), willistoni (Ritchie & Gleason, 1995), quinaria (Neems et al., 1997) и obscura (Noor & Aquadro, 1998) групп.
Межвидовые различия в Drosophila брачных звуках характеризуются одним или несколькими звуковыми параметрами, такими как: интерпульсовый интервал (IPI), который представляет собой время от начала одного импульса до начала следующего; интервал внутри импульса (PI), который является продолжительностью импульса; частота внутри импульса (IF), которая является акустически самой сильной частотой внутри импульса; количество импульсов в серии; и продолжительность каждого интервала внутри серии (Hoikkala & Lumme, 1987; Wheeler и др., 1988). Хотя еще не установлено, какие звуковые параметры обеспечивают специфичность, есть доказательства того, что IPI важен. Согласно Ritchie & Kyriacou (1996), IPI брачных звуков Drosophila варьируется очень мало в пределах естественных популяций и важен для репродуктивного успеха самцов.
Тем не менее, очевидно, что многие виды Drosophila производят сложные звуки с различными характеристиками, одна или несколько из которых могут быть полезны для идентификации видов. Эта специфичность в брачных звуках Drosophila может быть объяснена отбором на различение между видами (Tomaru & Oguma, 1994) различных типов звуков, которые могли эволюционировать через интраспецифический сексуальный отбор (Ritchie & Gleason, 1995). Кроме того, различные типы звуков, найденные в различных видах Drosophila, могут просто отражать филогенетические различия.
Подгруппа фасциола
Виды подгруппы fasciola встречаются во влажных лесах, в основном в Центральной и Южной Америке, а также на Карибских островах. В Бразилии их собирали в Атлантическом тропическом лесу, а также в лесах штатов Сан-Паулу, Парана, Мату-Гросу-ду-Сул и Баия (Вилела, 1983; Вилела и др., 1983; Тидон-Склорз и Сене, 1992).
Большинство исследований этой подгруппы было цитологическим и проводилось в начале 60-х годов, показывая, что виды в этой группе отличаются от всех остальных видов группы repleta наличием гомозиготных инверсий: 2o2, 2e3 и 2l3. Уассерман (1982) предложил общее происхождение для подгрупп fasciola и mulleri, поскольку у них есть общая инверсия 3c. Однако недавно Диниз (1998) показал, что инверсия 3c подгруппы fasciola отличается от той, что найдена в подгруппе mulleri, и предложил, что подгруппа fasciola произошла непосредственно от примитивной I и больше не может считаться связанной с подгруппой mulleri. Кун и др. (1995) провели кариотипическое исследование некоторых видов подгруппы fasciola и обнаружили межвидовую вариацию в размере микрохромосомы и в числе хромосом.
Согласно Вассерману (1992), подгруппа fasciola состоит из девяти видов: D. pictilis и D. pictura, образующих комплекс pictilis; D. paraguttata; D. mojuoides и D. moju, образующих комплекс moju; и D. fulvalineata, D. fasciola, D. coroica и D. fascioloides, которые не входят ни в один комплекс. Вилела (1983), изучая морфологию мужских половых органов, установил, что вид D. fascioloides, изученный Вассерманом (1962), на самом деле не принадлежит этому виду, и переименовал его в D. ellisoni. Он также добавил виды D. linearepleta и D. onca в подгруппу fasciola и описал D. carolinae, D. hermionae, D. ivai, D. querubimae, D. rosinae и D. senei как новые виды, увеличив количество видов в этой подгруппе до 18.
В этой работе мы описываем звуковые паттерны ухаживания шести видов подгруппы fasciola: D. ellisoni, D. fascioloides, D. moju, D. onca, D. rosinae и D. coroica. Кроме межвидовых анализов, мы также изучили различия в звуковых паттернах трех различных популяций D. coroica.
Материалы и методы
Создание и поддержание изолятов
Изоляторы были получены от отдельных самок, уже спарившихся в дикой природе. Эти изоляторы были идентифицированы с помощью морфологического анализа половых органов самцов F1 (Vilela, 1983). Личинки третьей инстанции подгруппы fasciola обычно пытаются выбраться из культуры. Чтобы избежать этой проблемы, культуры были открыты и помещены на влажный песок внутри пластиковых бутылок, чтобы личинки могли окуклиться в песке. Выведенные мухи были удалены с помощью аспиратора и перенесены в пробирки, заполненные питательной средой, где они созревали, спаривались и откладывали яйца, вновь начиная цикл.
Запись звуков
Недавно выведенные девственные мухи были изолированы в 35 мл пробирках и оставлены на 7-10 дней, чтобы дать время для полового созревания. Затем в камеру для записи звука, описанную Sene & Manfrin (1998), были помещены самец и самка одного и того же происхождения. Для записи звука камера с парой мух была помещена на диафрагму ультрачувствительного микрофона, как описано Manfrin и др. (1997). Мухи наблюдали в течение максимум 10 минут или до спаривания. Все записи проводились при температуре 25 ± 1ºC.
Звуковой анализ
Электромагнитные сигналы аналоговых записей были преобразованы в цифровые сигналы с частотой 5 кГц с помощью аналогово-цифрового преобразователя CAD 12/36-60 k (Lynx) и программы AqDADOS 4 (Lynx).
Звуковые паттерны ухания шести различных видов подгруппы fasciola описаны в Таблице 1. Также были проанализированы звуковые паттерны самцов D. coroica из трех различных мест для изучения межпопуляционных различий.
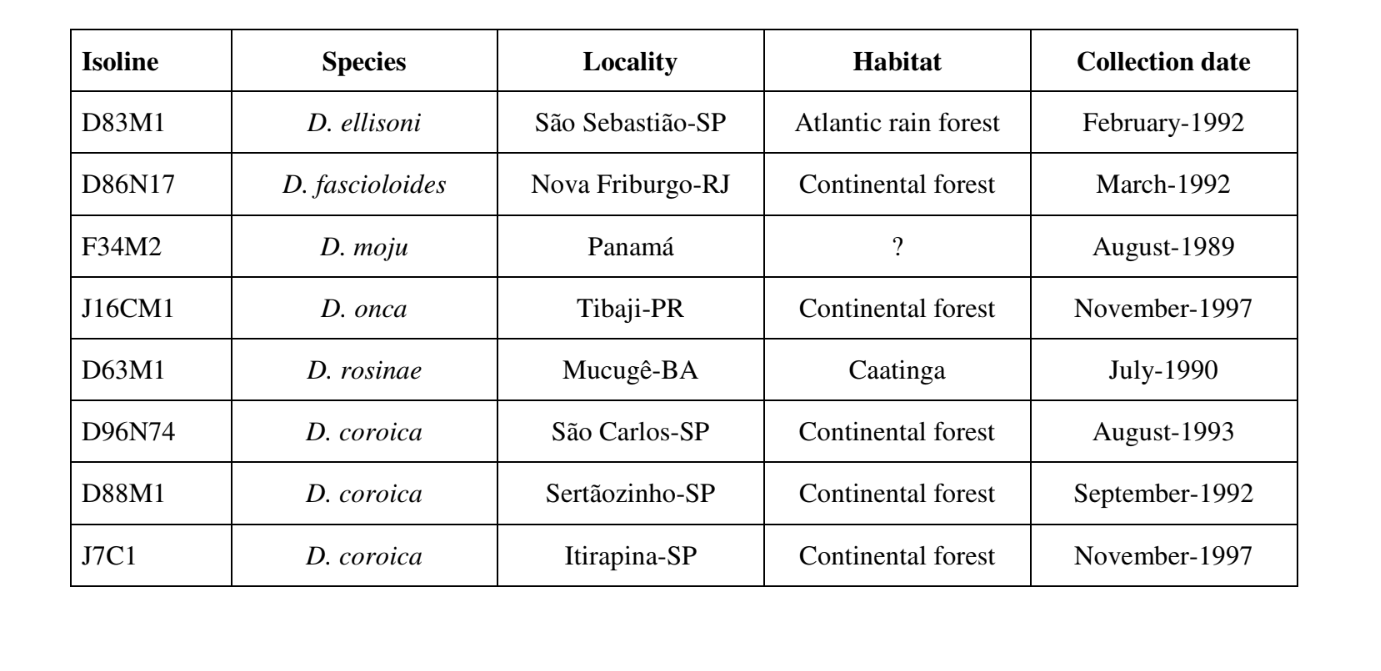
Параметры, использованные для сравнительных исследований звуковых паттернов, включали интервал внутри импульса (PI), интервал между импульсами (IPI) и частоту внутри импульса (IF). Значения PI и IPI были получены непосредственно из фигур. Значения IF были получены с помощью программного обеспечения MATLAB (MathWorks, Inc.) с использованием быстрого преобразования Фурье (FFT). Этот анализ разбивает звуковой сигнал на серию волн (синусоид и косинусоид) различных частот и амплитуд. IF звука определяется как частота волны с наибольшей амплитудой в звуковом сигнале.
FFT может анализировать звуки, содержащие тысячи точек за миллисекунду, но максимальная эффективность анализа достигается путем ограничения числа точек до степеней двойки. По этой причине все интервалы, использованные для анализа IF, были выбраны так, чтобы они имели количество точек, равное степеням двойки. Поэтому для D. moju использовались интервалы по 64 точки; для D. rosinae и D. coroica — 128 точек; для D. ellissoni — 256 точек; и для D. onca и D. fascioloides — 512 точек. Для каждого из 10 самцов из каждой из изолиний было сделано десять измерений параметров PI, IPI и IF.
Статистический анализ
Линии, описанные в Таблице 1, были проанализированы с целью исследования межвидовых различий. Изолинейка D96N74 была выбрана в качестве представителя D. coroica. Три линии (D96N74, D88M1 и J7C1) D. coroica были протестированы отдельно на предмет межпопуляционных различий. Значения параметров интерпульсного интервала (PI), интерпульсного интервала (IPI) и частоты внутри импульса (IF) были проанализированы отдельно с помощью дисперсионного анализа (ONE-WAY ANOVA) на уровне значимости 0.05.
Результаты
Рисунки 1-6 показывают основные сонограммы различных анализируемых видов. Изолинейка D96N74 была выбрана для представления вида D. coroica. Эти сонограммы и анализ звуковых импульсов позволяют разделить звуки ухания на три четкие группы: 1. звуки ухания видов D. onca, D. fascioloides и D. ellisoni, в которых каждая звуковая последовательность состоит из последовательности коротких импульсов, за которыми следует трель, аналогичная той, что встречается у некоторых видов подгруппы planitibia, обозначенной Хойккалой и др. (1994) как фразовая песня. У видов D. fascioloides и D. onca последовательность импульсов состоит из четырех импульсов, за которыми следует относительно длинная трель, в то время как у D. ellisoni всего два импульса, за которыми следует более короткая трель; 2. типичная для сонограммы D. rosinae, с уханием, образованным простыми, но дублированными импульсами, шаблон, также наблюдаемый Эуингом и Мияном (1986) у некоторых видов группы repleta; 3. тип, основанный на уханиях видов D. moju и D. coroica, с короткими импульсами в последовательности, аналогичными импульсной песне D. melanogaster (Шильхер, 1976) и звуку A D. mercatorum (Икеда и Маруо, 1982; Манфрин и др., 1997).
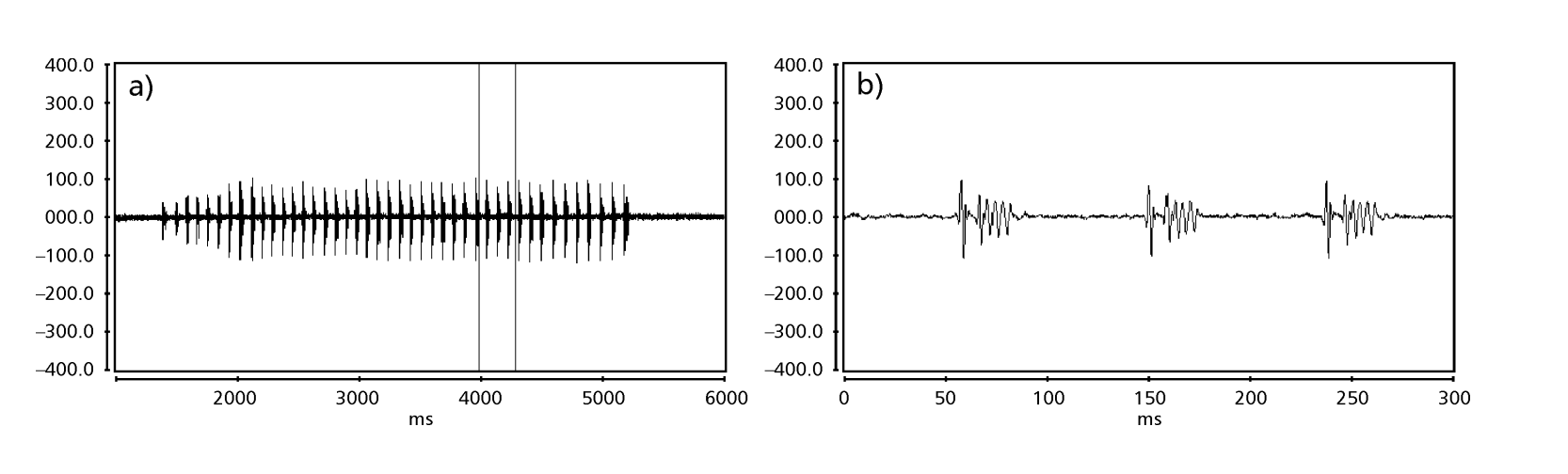
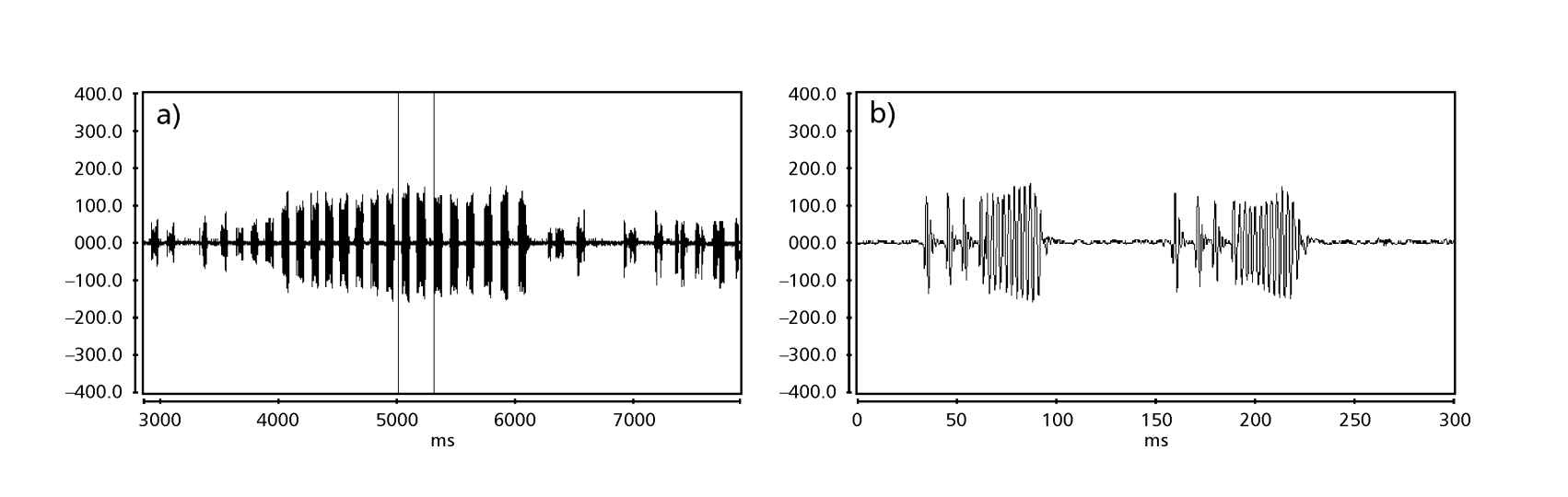
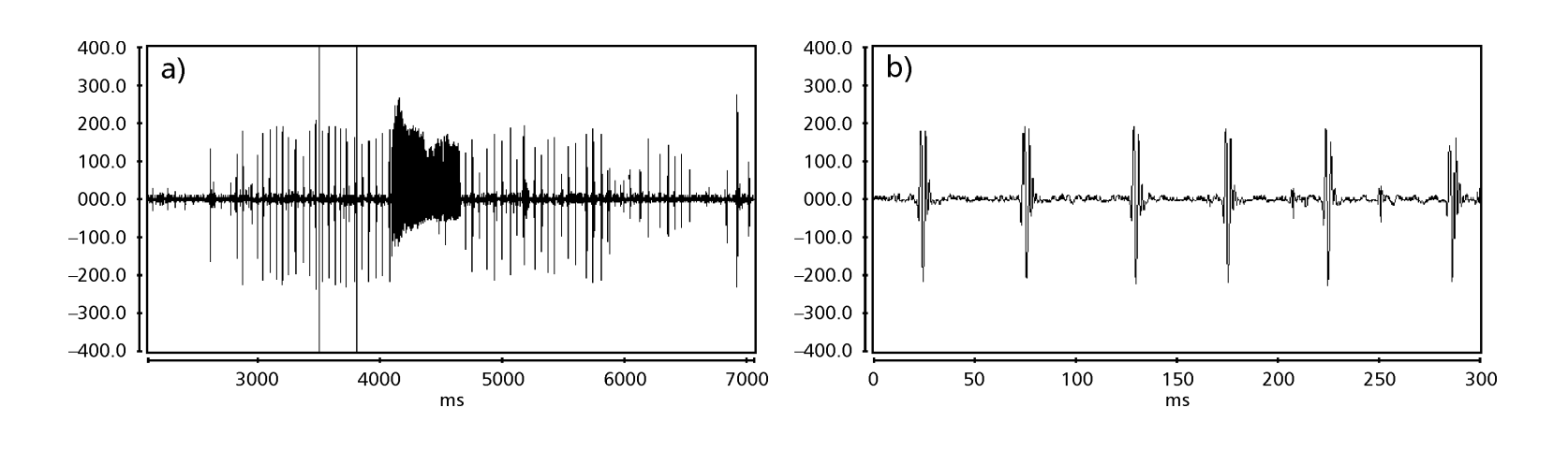
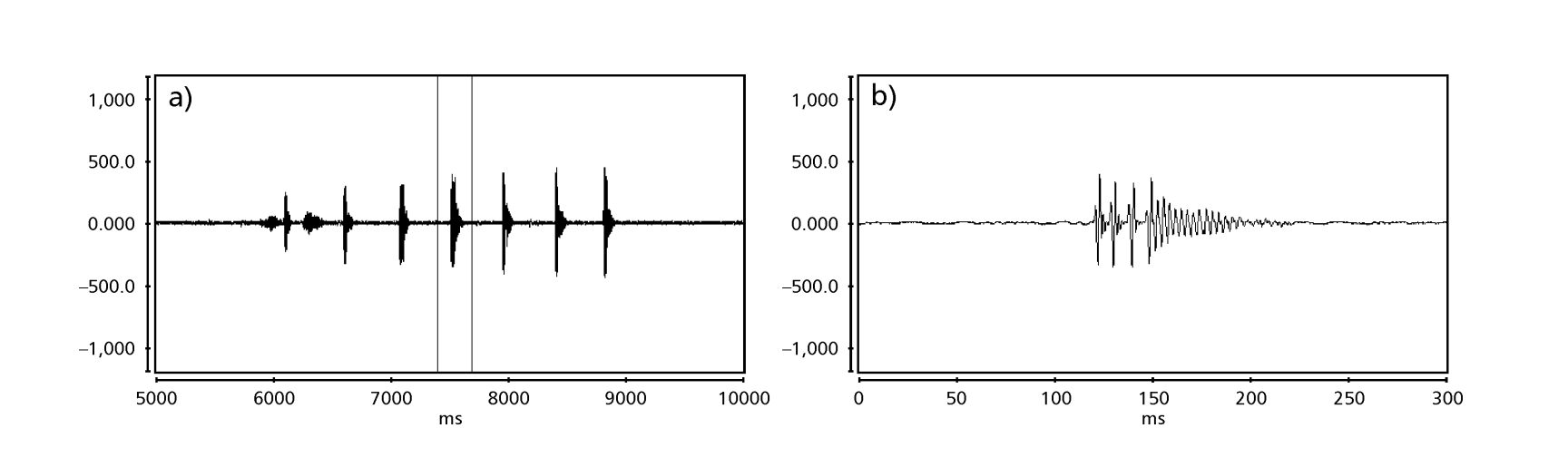
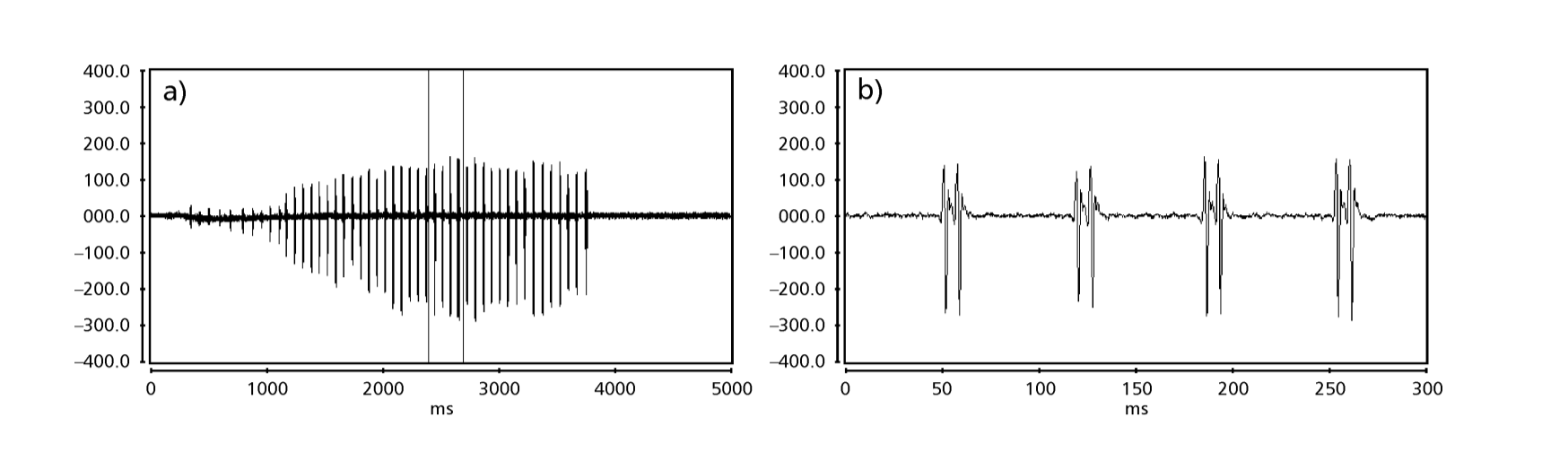
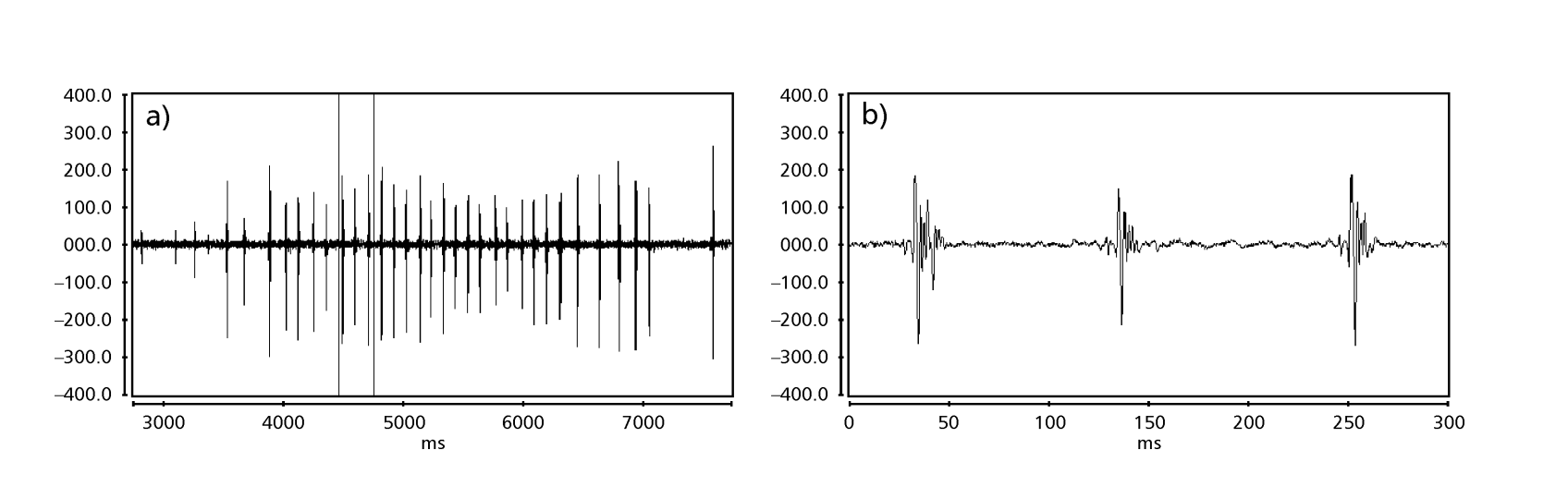
Сонограммы каждого из проанализированных видов представляли собой последовательность одного типа звукового импульса. Эта находка отличается от результатов исследований, проведенных на видах D. melanogaster (Schilcher, 1976), D. mercatorum (Ikeda & Maruo, 1982; Manfrin et al., 1997) и некоторых видов подгруппы repleta (Ewing & Miyan, 1986), в которых были обнаружены два разных типа звуков.
Таблица 2 показывает средние значения для PI (интервал внутри импульса), IPI (интервал между импульсами) и IF (внутриимпульсная частота) изолиний, описанных в Таблице 1. Сравнение стандартных отклонений (± sd) среди различных видов показывает, что D. ellisoni и D. fascioloides имеют наименьшую вариацию по всем трем параметрам: PI (27.470 ± 2.73 и 69.859 ± 6.12), IPI (97.473 ± 5.76 и 125.757 ± 7.87) и IF (251.358 ± 19.36 и 362.901 ± 17.04), что указывает на то, что звуки ухаживания этих видов однородны с небольшой внутривидовой вариацией. С другой стороны, D. onca имел значительную вариацию в PI (93.662 ± 31.44) и IPI (508.091 ± 73.06) и умеренно высокую вариацию в IF (289.260 ± 32.71) по сравнению с другими видами, что указывает на то, что различия в PI и IPI, хотя и большие, не были достаточными для значительного изменения формы импульсной волны. Стандартные отклонения были относительно низкими для PI (7.363 ± 7.36 и 14.673 ± 1.98) и IPI (50.859 ± 5.47 и 72.340 ± 5.68) в D. moju и D. rosinae, но значения IF были относительно высокими (412.501 ± 55.63 и 306.624 ± 57.80). В этом случае, хотя продолжительность импульса и время от одного импульса к следующему были относительно однородными внутри каждого из этих видов, форма звукового импульса значительно варьировала.
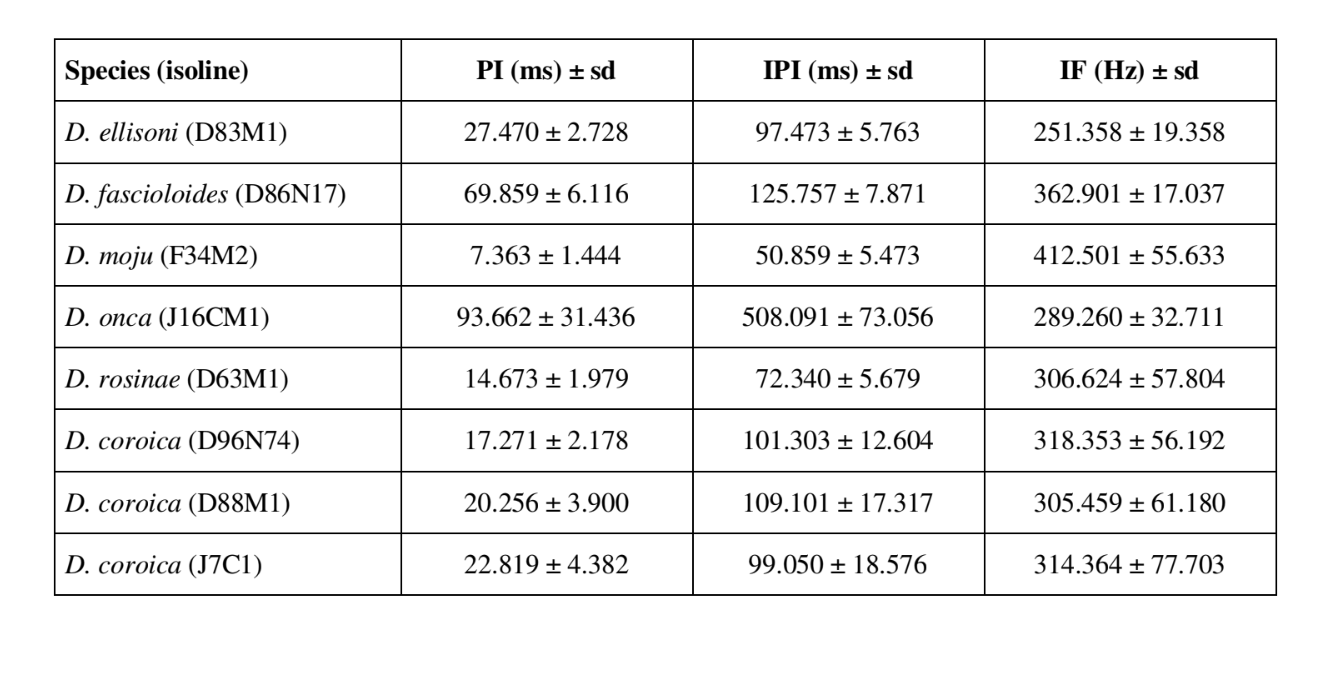
В пределах популяции было относительно мало вариаций в ПИ линий D. coroica D96N74, D88M1 и J7C1 (17.271 ± 2.18, 20.256 ± 3.90 и 22.819 ± 4.38 соответственно), промежуточная изменчивость в ИПИ (101.303 ± 12.60, 109.101 ± 17.32 и 99.050 ± 18.58) и высокая изменчивость в ИФ (318.353 ± 56.19, 305.459 ± 61.18 и 314.364 ± 77.70), что указывает на то, что даже при низкой внутрипопуляционной изменчивости в длине звукового импульса, существовала значительная изменчивость в форме звукового импульса.
Когда средние значения параметров ПИ и ИПИ были сравнены, различия среди видов подгруппы fasciola могли быть найдены хотя бы для одного из этих параметров. Эти различия в звуках ухаживания можно увидеть на рис. 7, где значения для этих двух параметров представлены в виде эллипсов, которые включают 75% значений для параметров ПИ и ИПИ. Значительные различия были найдены в значениях ПИ (ANOVA, F5,48 = 124.98, p < 0.001) и ИПИ (ANOVA, F5,48 = 949.35, p < 0.001) среди видов. Межвидовые различия в ИФ показаны на рис. 8. Значительные различия также были найдены в значениях ИФ (ANOVA, F5,48 = 23.40, p < 0.001).
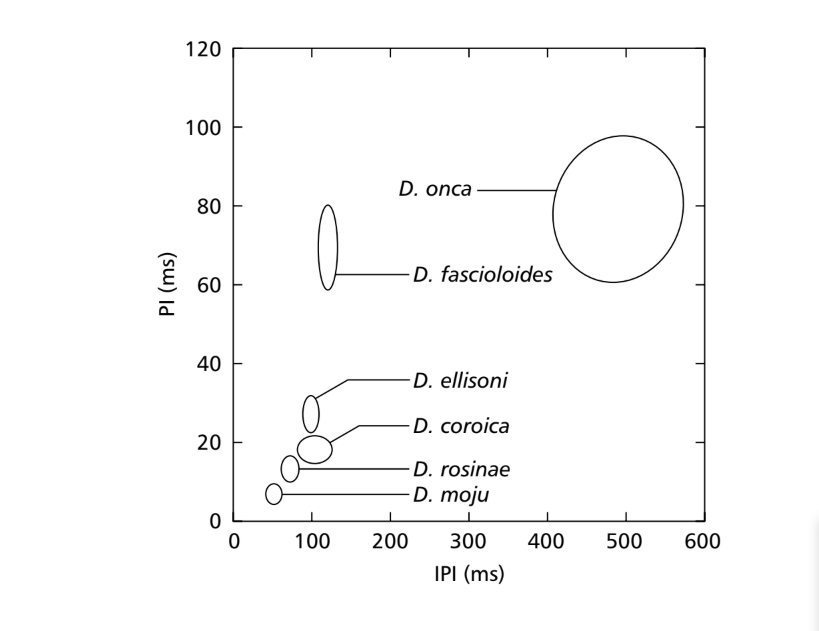
Три популяции D. coroica значительно различались по PI (ANOVA, F2,24 = 8.75, p < 0.05), но не по IPI (ANOVA, F2,24 = 1.72, p > 0.10) и IF (ANOVA, F2,24 = 0.15, p > 0.25).
Обсуждение
Хойккала и др. (1994) в своем анализе брачного звука различных видов подгруппы planitibia обнаружили, что общим является наличие одних и тех же типов звуков у филогенетически близких видов. Также Хойккала и Канеширо (1993) наблюдали, что некоторые близкородственные виды в этой подгруппе, которые являются симпатричными, используют разные типы звуков в брачных ритуалах.
Нур и Аквадро (1998) обнаружили значительные различия в брачных звуках у двух близкородственных видов подгруппы obscura: D. pseudoobscura и D. persimilis. Некоторые эксперименты с перекрестками между этими двумя видами показали, что самки D. pseudoobscura вероятно увеличили свою дискриминацию против самцов D. persimilis в симпатричных популяциях. Это могло произойти через отбор против производства плохо адаптированных гибридов, что привело к изменению брачного звука в этих симпатричных популяциях.
Эуинг и Миян (1986), работая с видами группы repleta, не нашли связи между филогенией, основанной на цитологических данных, и другой, составленной на основе звуковых данных. Тем не менее, некоторые характеристики, которые имеют большинство изученных видов и которые также встречаются у видов других групп, позволяют нам предложить предковый брачный звук для группы repleta. Этот звук, вероятно, состоял из двух различных компонентов: начального звука A, состоящего из максимум 10 коротких и регулярных моноциклических импульсов с интервалом между импульсами от 8 до 15 мс; и звука B, состоящего из серии регулярных и моноциклических импульсов, но с большим количеством импульсов и более длительным интервалом между импульсами в конце брачного периода. В ходе эволюции группы repleta некоторые виды могли утратить звук A, а другие — звук B, и у многих звук B стал менее регулярным и более сложным.
Основываясь на этих предположениях и наблюдая за сонограммами, полученными для шести видов подсгруппы fasciola в этом исследовании, мы можем сделать вывод, что у всех этих видов произошла утрата звука A, так как интервалы между импульсами всегда значительно превышают 15 мс. D. coroica имеет нерегулярные импульсы, со средним интервалом между импульсами 101,3 мс, в то время как брачные звуки D. moju состоят из более регулярных импульсов, со средним интервалом между импульсами
50.9 мс. D. rosinae имеет сонограмму, состоящую из дублированных импульсов, тип звука, также найденный Эуингом и Мияном (1986), со средним интервалом между импульсами 72.3 мс. D. ellisoni, D. fascioloides и D. onca имеют звуки ухаживания, состоящие из полициклических импульсов, вероятно, в результате слияния звуков A и B, со средними интервалами между импульсами 97.4, 125.7 и 508.0 мс соответственно. Эти сходства в типах звуков у этих трех видов не имеют тесной корреляции с цитологическими данными, за исключением D. ellisoni и D. fascioloides, которые были расположены близко друг к другу в филогении, основанной на цитологических признаках, у них есть два общих слияния хромосом: 2-4F и 3-5F (Диниз, 1998). Как и в случае морфологических признаков, ожидается тенденция к тому, что близкородственные виды будут разделять некоторые параметры звука. Тем не менее, даже когда филогенетические отношения между видами известны, эволюционные тенденции, основанные на звуковых признаках, не были хорошо установлены.
Хотя звук ухаживания не является примером циркадного поведения, исследования связали это поведение с экспрессией гена period (per), расположенного на X-хромосоме и хорошо охарактеризованного на молекулярном уровне. Мутации в этом гене могут изменять циркадный ритм Drosophila. Генетический анализ, проведенный Конопкой и др. (1996), показал, что звуковой ритм интерпульсного интервала также зависит от мутаций в гене per. Кириакоу и Холл (1980) сообщили о ритмической модуляции интерпульсного интервала с определенными периодами у D. melanogaster и D. simulans соответственно 55 и 35 секунд. Эти различия были показаны как связанные с некоторыми вариациями в гене per. Мутации в этом гене, которые изначально изучались Конопкой и Бенцером (1971) в анализе аномальных циркадных ритмов, вызывали соответствующие эффекты в ритмах интерпульсного интервала звуков ухаживания. Мутация perShort (perS), с циркадным ритмом около 19-20 часов, имела средний период 41.5 секунды; мутация perLong (perL), чей циркадный ритм составлял 28-30 часов, имела средний период 82.1 секунды; и мутация per0 не имела ритма интерпульсного интервала.
В этом исследовании мы обнаружили значительные различия в параметрах интерпульсного интервала, интерпульсного интервала и частоты внутри импульса в анализируемых видах fasciola. Среди этих параметров интерпульсный интервал оказался тем, который отличался больше всего, с небольшой суперпозицией и малой внутривидовой вариацией, что предполагает, что этот параметр может быть частью системы распознавания видов. Этот высокий уровень дифференциации в звуках самцов согласуется с общим мнением о том, что брачное поведение может отличаться больше, чем морфологические характеристики или другие особенности у родственных видов. Однако в данный момент невозможно узнать направление эволюции этих различных типов звуков в подгруппе fasciola. Генетические исследования показали, что межвидовые различия в интерпульсных интервалах часто обусловлены аддитивными и полигенными факторами. Эти различия также могут быть объяснены адаптацией к различным условиям окружающей среды в отсутствие генетического потока. Известно, что в четвертичный период имели место чередующиеся циклы между холодно-сухим и жарко-влажным климатом. В цикле холодно-сухого климата растительность, адаптированная к сухим условиям, расширялась, в то время как леса сокращались. Во время последующего жарко-влажного цикла ситуация менялась на противоположную (Ab’ Saber, 1977; Vanzolini, 1981). Считается, что мухи следовали за этими процессами сокращения и расширения лесов, что приводило к длительным периодам географической изоляции во время холодно-сухих циклов и некоторым областям симпатри во время жарко-влажных циклов.
Хотя интервал между импульсами может значительно варьироваться среди различных видов рода Drosophila, обычно это не наблюдается при анализе популяций одного и того же вида (Costa и др., 2000; Ritchie и др., 1994). Три проанализированные популяции D. coroica имели значительные различия только в интервале внутри импульса, без различий в интервале между импульсами и частоте внутри импульса. Возможно, что на интервал между импульсами и частоту внутри импульса действует сильное селекционное давление. Эта низкая изменчивость также может возникнуть, если географически различные популяции были генетически однородными из-за постоянного генного потока или если общий предок недавно колонизировал эти популяции. Однако данные PI показывают, что это не так.
Мало что известно о генетических механизмах, участвующих в процессах видообразования, однако сначала необходимо понять генетику и эволюцию механизмов, препятствующих генному потоку. Бесплодные и нестабильные гибриды считаются сильными барьерами для генного потока, но такие постзиготные механизмы могут привести к высоким затратам в виде гамет и энергии. Поэтому различия в звуках ухаживания могут быть важным изолирующим механизмом, так как они ограничивают или предотвращают такие потери (Welbergen и др., 1992). Анализ этих сигналов, которые важны для успеха спаривания, представляет значительный интерес для эволюционной биологии, и знание архитектуры звуков может помочь подтвердить различные модели сексуального отбора и видообразования.
Авторы: Коста, К. Т. А. и Сене, Ф. М.
Ссылки:
- Аб’ Сабер, А. Н., 1977, Пространства, занятые расширением сухих климатов Южной Америки, во время четвертичных ледниковых периодов. Палеоклиматы, 3: 1-19.
- Чанг, Х. Ц. и Миллер, Д. Д., 1978, Звуки ухаживания и спаривания у видов подгруппы Drosophila affinis. Эволюция, 32: 540-550.
- Коста, К. Т. А., Кун, Г. Ц. С. и Сене, Ф. М., 2000, Низкая вариация ухаживающей песни в южных и юго-восточных бразильских популяциях D. meridionalis (Двукрылые, Drosophilidae). Rev. Brasil. Biol., 60: 53-61.
- Коулинг, Д. Е. и Бернет, Б., 1981, Ухаживающие песни и генетический контроль акустических характеристик у родственных видов подгруппы Drosophila melanogaster. Животное поведение, 29: 924-935.
- Диниз, Н. М., 1998, Хромосомная филогения видов подгруппы fasciola группы repleta рода Drosophila. Диссертация на соискание ученой степени, Медицинский факультет Университета Рибейран-Прету, Университет Сан-Паулу.
- Эуинг, А. В. и Миян, Дж. А., 1986, Половой отбор, сексуальная изоляция и эволюция песни в группе видов Drosophila repleta. Животное поведение, 34: 421-429.
- Хойккала, А. и Лумме, Х., 1987, Генетическая основа эволюции мужских ухаживающих звуков группы Drosophila virilis. Эволюция, 41: 827-845.
- Хойккала, А. и Канэширо, К. Я., 1993, Изменение последовательности сигнал-ответ, ответственной за асимметричную изоляцию между Drosophila planitibia и D. silvestris. Proc. Natn,. Acad. Sci., 90: 5813-5817.
- Хойккала, А. и Исоехранен, Е., 1997, Вариация и повторяемость характеристик ухаживающей песни среди диких и лабораторно выведенных самцов Drosophila montana и D. littoralis (Двукрылые-Drosophilidae). Журнал поведения насекомых, 10: 193-202.
- Хойккала, А., Лаковаара, С. и Ромппайнен, Е., 1982, Поведение при спаривании и ухаживающие звуки самцов группы Drosophila virilis. В: Лаковаара, С. (ред.), Достижения в генетике, развитии и эволюции Drosophila. Пленум, Нью-Йорк.
- Хойккала, А., Кеннет, Я. и Хой, Р. Х., 1994, Ухаживающие песни видов подгруппы с картинными крыльями Drosophila planitibia. Животное поведение, 47: 1363-1374.
- Икеда, Х. и Маруо, О., 1982, Направленный отбор по частоте повторения импульсов ухаживающего звука и связанные с ним изменения в нескольких характеристиках у Drosophila mercatorum. Японский журнал генетики, 57: 241-258.
- Конопка, Р. Дж. и Бензер, С., 1971, Мутанты часов у Drosophila melanogaster. Proc. Natl. Acad. Sci., 68: 2112-2116.
- Конопка, Р. Дж., Кириаку, С. П. и Холл, Дж. С., 1996, Мозаичный анализ в ЦНС Drosophila циркадных ритмов ухаживающей песни, затронутых мутацией часов. Ж. Нейрогенет., 11: 117-140.
- Кун, Г. Ц. С., Диниз, Н. М., Алвеш, М. А. Р. и Сене, Ф. М., 1995, Хромосомная конституция видов подгруппы fasciola группы repleta рода Drosophila. Rev. Bras. Gen., 18 supplement: 282.
- Кириаку, С. П. и Холл, Дж. С., 1980, Мутации циркадного ритма в Drosophila melanogaster влияют на краткосрочные колебания в ухаживающей песне самца. Proc. Natl. Acad. Sci., 77: 6729-6733.
- Манфрин, М. Х., Прадо, П. Р. Р. и СЕНЕ, Ф. М., 1997, Анализ звуковых компонентов сексуального ухаживания двух подс Species Drosophila mercatorum (Двукрылые, Drosophilidae). Rev. Brasil. Biol., 57: 349-355.
- Нимс, Р. М., Духер, К., Батлин, Р. К. и Шоррокс, Б., 1997, Различия в ухаживающей песне самцов среди видов группы quinaria рода Drosophila. Журнал поведения насекомых, 10: 237-246.
- Нур, М. А. и Аквадро, К. Ф., 1998, Ухаживающие песни Drosophila pseudoobscura и D. persimilis: анализ вариации. Животное поведение, 56: 115-125.
- Ритчи, М. Г. и Глисон, Дж. М., 1995, Быстрая эволюция паттерна ухаживающей песни у видов Drosophila willistoni. Ж. Эвол. Биол., 8: 463-479.
- Ритчи, М. Г. и Кириаку, С. П., 1996, Искусственный отбор на сигнал ухаживания у Drosophila melanogaster. Животное поведение, 52: 603-611.
- Ритчи, М. Г., Яте, В. Х. и Кириаку, С. П., 1994, Генетическая изменчивость интервала между импульсами ухаживающей песни среди некоторых европейских популяций Drosophila melanogaster. Наследственность, 72: 459-464.
- Шильхер, Ф. В., 1976, Роль слуховых стимулов в ухаживании Drosophila melanogaster. Животное поведение, 24: 18-26.
- Сене, Ф. М. и Манфрин, М. Х., 1998, Камера для записи ухаживающей песни у Drosophila. D. I. S., 81: 203-204.
- Шорей, Х. Х., 1962, Природа звука, производимого Drosophila melanogaster во время ухаживания. Наука, 137: 677-678.
- Тидон-Склорз, Р. и Сене, Ф. М., 1992, Вертикальное и временное распределение видов Drosophila (Двукрылые, Drosophilidae) в лесной зоне штата Сан-Паулу, Бразилия. Rev. Brasil. Biol., 52: 311-317.
- Томару, М. и Огума, Я., 1994, Различия в ухаживающей песне у видов комплекса Drosophila auraria. Животное поведение, 47: 133-140.
- Ванзолини, П. Е., 1981, Квази-исторический подход к естественной истории дифференциации рептилий в тропических географических изолятах. Папейш Авулсос Зоол., 34: 189.
- Вилела, К. Р., 1983, Ревизия группы видов Drosophila repleta (Двукрылые, Drosophilidae). Revta. Bras. Ent., 27: 1-114.
- Вилела, К. Р., Перейра, М. А. Q. Р. и Сене, Ф. М., 1983, Предварительные данные о географическом распределении видов Drosophila в морфоклиматических доменах Бразилии. II. Группа repleta. Cienc. Cult., 35: 66-70.
- Уассерман, М., 1962, Цитологические исследования группы repleta рода Drosophila. VI. Подгруппа fasciola. Университет Техаса Паблик., 6205: 119-134.
- Уассерман, М., 1982, Эволюция группы repleta. В: М. Эшбернер, Х. Л. Карсон и Дж. Н. Томпсон (ред.), Генетика и биология Drosophila, т. 3b. Академическое издательство, Лондон, стр. 61-139.
- Уассерман, М., 1992, Цитологическая эволюция группы видов Drosophila repleta: VI. Подгруппа fasciola. В: Исследования в генетике II. Исследовательские отчеты по генетике Drosophila, таксономии и эволюции. CRC Press, Inc., М. Р. Университет Техаса, Остин.
- Велберген, П., Ван Дюкен, Ф. Р., Шарлоо, В. и Кёлер, В., 1992, Генетическая основа сексуальной изоляции между Drosophila melanogaster и D. simulans. Эволюция, 46: 1385-1398.
- Уилер, Д. А., Филдс, В. Л. и Холл, Дж. С., 1988, Спектральный анализ ухаживающих песен Drosophila: D. melanogaster, D. simulans и их межвидовой гибрид. Поведенческая генетика, 18: 675-703.